

Los estudios epidemiológicos llevan generalmente a la conclusión que el consumo moderado de bebidas alcohólicas protege contra las enfermedades cardiovasculares (ECV) en comparación a los sujetos abstemios y a los bebedores en exceso (Rimm et al., 1996). Los datos actuales permiten avanzar que las diferentes bebidas alcohólicas no son equivalentes entre ellas, y que el vino podría proporcionar una mayor protección específica frente a las ECV (Gronbaek et al., 1995; Renaud et al., 1999). Los polifenoles (polifenoles) que contiene, variables en clase y cantidad según el tipo de vino considerado, son sustancias producidas por el metabolismo secundario de la vid. Su estudio suscita un interés creciente a causa de sus propiedades antioxidantes – potencialmente antiaterogénicas si nos basamos en la teoría oxidativa de la aterosclerosis – y de sus efectos sobre la mediación celular (Léger et al., 2000a). En el estudio de los efectos de los polifenoles del vino sobre la salud se debe tener en cuenta si se encuentran a disposición de las células (biodisponibilidad), y la aparición de derivados metabólicos después de su ingestión. A pesar de las reservas que puedan surgir al considerar la equivalencia de las nociones de biodisponibilidad y concentración plasmática, se puede hacer una aproximación práctica a la biodisponibilidad de los polifenoles evaluando las concentraciones plasmáticas de los componentes procedentes del vino que se encuentran presentes en la sangre.
Resultados y discusión
Hemos observado recientemente que el pico de catequina plasmática libre y glucuronidada después de una toma de vino tinto (300 mL dentro de una comida que aporte unas 500 Kcal, sin contar el alcohol) sería, en determinados sujetos, del orden de 0,1 µmol/L (valor máximo) (Léger et al., 2000a), y hemos demostrado que la capacidad antioxidante del plasma es máxima coincidiendo con el pico de catequina, es decir, 3-4 h después de la comida. La concentración de catequina en ayunas se encuentra bajo el límite actual de detección electroquímica, inferior a 0,02 µmol/L, y en lo referente a los ácidos gálico y cafeic, la concentración plasmática es inferior a 0,01 µmol/L. En las mismas condiciones, y con un sistema de detección por espectrometría de masas, se han descrito en ayunas valores de catequina libre, glucuronidada, metilada y sulfatada inferiores a 0,002 µmol/L, y valores comparables a los nuestros después de una toma de vino tinto (Donovan et al., 1999). En general, si se desea seguir las variaciones en el contenido de polifenoles en la sangre, el conjunto de datos actuales indica claramente que el límite de detección debe disminuir por lo menos en un factor de 100. Para responder a esta preocupación, en nuestro laboratorio se están ensayando derivatizaciones químicas de cara al análisis y la cuantificación de la catequina y los ácidos fenólicos por cromatografía – espectrometría de masas.
Por otro lado hemos mostrado que los polifenoles del vino tinto se adsorben a la superficie de las lipoproteínas después de su paso a través de la pared intestinal, y que son, al menos en parte, transportados asociados a las LDL (Carbonneau et al., 1997; Carbonneau et al., 1998), de las que se pueden liberar fácilmente por simple diálisis o por cromatografía líquida de gel filtración. Creemos que este resultado se debe a la presencia de los polifenoles en la interfase aceite/agua, y no en el núcleo lipídico de la partícula de LDL, de acuerdo con su carácter esencialmente hidrosoluble. Por lo que respecta a la catequina y a los otros ácidos fenólicos, el ácido gálico es el que se encuentra más fuertemente asociado a las LDL (Cartron et al., 2001) como muestra la figura 1, hecho que podría tener consecuencias fisiológicas particulares en términos de transporte a los tejidos extrahepáticos.
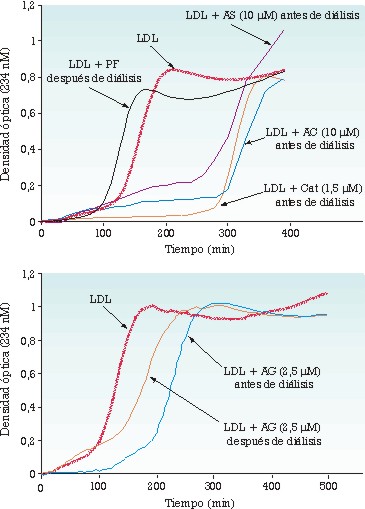
Hemos clasificado los polifenoles en función de su actividad antioxidante específica (AAE, actividad referida a la concentración de sustancia activa) frente a las LDL (Cartron et al., 2001). Es interesante destacar que la AAE de los ácidos benzoicos (gálico y protocatéquico) siempre es más elevada que la de la catequina y el ácido cafeico, y que la AAE aumenta a medida que disminuye la concentración de los ácidos benzoicos, condición que no se cumple para los restantes polifenoles probados. El hecho de que se obtengan valores altos de AAE para concentraciones submicromolares de ácido gálico presentes en el plasma (véase más arriba) ha conducido a la hipótesis de que en condiciones fisiológicas este ácido fenólico actúa como antioxidante.
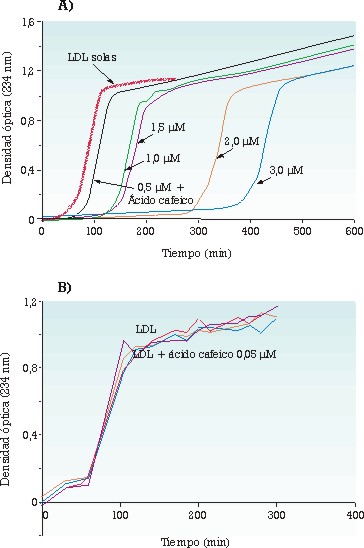
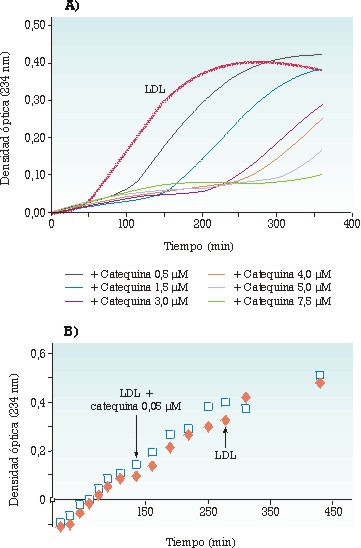
Se han utilizado dos sistemas in vitro para medir la AAE frente a las LDL: la lipoperoxidación por cobre (de 50/1 a 100/1, Cu2+/LDL, mol/mol) y la lipoperoxidación producida por descomposición térmica del 2,2’-nitro bis(2-amidinopropano) (25/1, NAPH/LDL, mol/mol), que generan radicales libres oxigenados orgánicos (alcoxi/alcoperoxi). La catequina presenta una AAE más alta en el sistema NAPH que en el sistema Cu2+, mientras que el inverso se observa para el ácido gálico. El ácido cafeic presenta una AAE similar en los dos sistemas. Estos resultados subrayan el papel acomplejante del ácido gálico frente al Cu2+ (o su interacción sobre el lugar de unión del Cu2+ a las LDL), y la principal función de la catequina como trampa de radicales libres. Estos dos sistemas han permitido igualmente mostrar que, en condiciones estándar (LDL 0,1 µmol/L), la resistencia a la oxidación de las LDL depende estrechamente de las concentraciones micromolares de ácido cafeico o de catequina, sea cual sea el sistema oxidante utilizado, y que esta resistencia no se ve modificada en condiciones próximas a las plasmáticas (LDL 1 µmol/L, catequina o ácido cafeico 0,05 µmol/L) (figuras 2 y 3). Los resultados obtenidos sugieren que los conocidos efectos protectores frente a la oxidación de estos dos compuestos fenólicos no son compatibles con sus concentraciones fisiológicas o con la concentración de las LDL presentes en la sangre circulante.
De la misma manera, hemos podido demostrar que la capacidad antioxidante de los polifenoles frente a las LDL depende del contenido en vitamina E de estas. Ello concuerda con la idea que la vitamina E funciona en el interior de la partícula lipoproteica como un dispositivo antioxidante liposoluble indispensable para la expresión de la actividad antioxidante de los polifenoles hidrosolubles.
Se ha examinado la respuesta celular a los polifenoles con la línea transfectada MCF-7, determinando la actividad transcripcional de la proteína 1 de activación (AP-1), y con la línea monocítica diferenciada THP-1, midiendo la producción de anión superóxido (O2•-). Los ácidos cafeico y gálico inhiben parcialmente ambas actividades celulares, aunque el ácido gálico lo hace a concentraciones menores que el cafeic. El hidroxitirosol, un polifenol presente en las aceitunas, el aceite de oliva y el alpechín, presenta el mismo efecto sobre la producción de O2•- (Léger et al., 2000b). Estas acciones podrían explicar los efectos supresores de las estructuras polifenólicas de tamaño pequeño sobre la proliferación y la diferenciación celulares (dado que la AP-1 está directamente implicada en estos fenómenos) y sobre la formación de las células espumosas (ya que el O2•- se encuentra directamente implicado en la formación de las LDL oxidadas, responsables de la sobrecarga lipídica de los macrófagos que los transforma en células espumosas), efectos que dan fe de un papel antiaterogénico. El ácido gálico actúa a concentraciones demasiado bajas para considerar una acción real a condiciones fisiológicas.
Conclusión
Los estudios in vitro registrados en la literatura determinan la acción de compuestos fenólicos presentes en el vino, a concentraciones muy superiores a las que se encuentran en el plasma (véase, por ejemplo, Andriambeloson et al., 1997). Las menores concentraciones utilizadas in vitro son diez veces superiores a las concentraciones plasmáticas más elevadas, encontradas en condiciones posprandiales, y son al menos 500 veces superiores a las concentraciones plasmáticas en ayunas. Los resultados que hemos presentado sugieren claramente que las concentraciones plasmáticas de los compuestos fenólicos del vino no explican el efecto fisiológico de protección frente a la oxidación o la mediación celular de estas sustancias, probablemente con excepción del ácido gálico. Será necesario en el futuro conocer mejor los derivados metabólicos producidos a partir de las moléculas parentales presentes en el vino, ya que podría ser que estos derivados tuvieran efectos biológicos superiores a los de sus precursores. Esta hipótesis permitiría reconciliar los resultados de los estudios in vitro con los obtenidos in/ex vivo (consultar los resultados sobre la capacidad antioxidante del plasma y la literatura citada [Léger et al., 2000a]) que describen un efecto antioxidante plasmático real del vino en condiciones fisiológicas.
Bibliografía
Andriambelosom E., Kleschyov A.L., Muller B., Beretz A., Stoclet J.C., and Andriantsitohaina R.: «Nitric oxyde production and endothelium-dependent vasorelaxation induced by wine polyphenols in rat aorta», British Journal of Pharmacology 1997; 120: 1053-1058.
Carbonneau M.A., Léger C.L., Descomps B., Senglat C., Fouret G., Michel F., Monnier L.: «Increase of antioxydant capacity of plasma and vitamin E content of LDL in human volunteers receiving red-wine alcohol-free phenolic compounds», Proceedings of the World Conference on Oilseed and Edible Oils Processing, Vol 2 : Advances in Oils and Fats, Antioxidants and Oilseed By-Products 1998. AOCS Press, Champaign, Ill, 284-288.
Carbonneau M.A., Léger C.L., Monnier L., Bonnet C., Michel F., Fouret G, Dedieu F. and Descomps B.: «Supplementation with red wine phenolics increases the antioxidant capacity of plasma and vitamin E of low density lipoprotein without changing the lipoprotein Cu2+– oxidability: Possible explanation by phenolics location», European Journal of Clinical Nutrution 1997; 51: 682-690.
Cartron E., Carbonneau M-A., Fouret G., Descomps B., Léger C.L.: «Specific antioxidant activity of caffeoyl derivatives and other natural phenolic compounds: LDL protection against oxidation and decrease in the proinflammatory lysophosphatidylcholine production», Journal of Natural Products 2001; 64(4):480-486.
Donovan J.L., Bell J.R., Kasim-Karakas S., Bruce German J., Walzem R.L., Hansen R.J. and Waterhouse A.L.: «Catechin is present as metabolites in human plasma after consumption of red wine», Journal of Nutrition 1999; 129: 1662-1668.
Léger C.L., Carbonneau M.A., Descomps B.: «Consommation de vin et prévention contre les maladies cardiovasculaires», Alimentation Méditerranéenne et Santé : actualités et perspectives, John Libbey Eurotext 2000a: 55-71.
Léger C.L., Kadiri-Hassani N., Descomps B.: «Decreased superoxide anion production in cultured human promonocyte cells (THP-1) due to polyphenol mixtures from olive oil processing wastewaters», Journal of Agriculture and Food Chemistry 2000b; 48: 5061-5067.
Renaud S.C., Gueguen R., Siest G., Salomon R.: «Wine, beer, and mortality in middle-aged men from eastern France», Archives of Internal Medicine 1999; 159: 1865-1869.
Rimm E.B., Klatsky A., Grobbee D., Stampfer M.J.: «Review of moderate alcohol consumption and reduced risk of coronary heart disease : is the effect due to beer, wine, or spirits ?», British Medical Journal 1996; 312: 731-736.



{kind=link}