

Uno de los principales objetivos en la elaboración de vino tinto es optimizar y resaltar, mediante la fermentación, el carácter frutal de la variedad y el carácter territorial de las uvas. Además de la uva, tanto las bacterias como las levaduras tienen un papel relevante a la hora de conferir un estilo al aroma y el sabor del vino. Se sabe que las levaduras contribuyen al aroma y sabor del vino mediante la fermentación alcohólica (Swiegers et al., 2005). No obstante, las bacterias a menudo se perciben solo como agentes de la conversión de ácido málico en láctico por medio de la fermentación maloláctica (FML), o por malograr el vino. Mientras que se acepta generalmente que la conversión ácida durante la FML da lugar a una suavización del vino gracias a un incremento del pH y una disminución de la acidez, cada vez se reconoce más que Oenococcus oeni puede modular el aroma del vino durante su crecimiento y el metabolismo de una larga lista de compuestos (Swiegers et al., 2005; Bartowsky y Pretorius, 2008). En particular, los resultados más recientes indican que la FML puede modular ciertos aromas volátiles asociados con propiedades sensoriales afrutadas en el vino. O. oeni es la principal especie de bacteria de ácido láctico asociada con la FML, ya que puede sobrevivir y prevalecer en el –a menudo– duro entorno enológico (pocos nutrientes, elevado contenido alcohólico y pH). Sin embargo, a pesar de que la población endógena natural de O. oeni puede crecer y conducir la FML, los resultados impredecibles asociados a una fermentación maloláctica espontánea han llevado al desarrollo de preparaciones bacterianas de cultivos iniciadores que contienen cepas específicas de O. oeni. La adición de estos cultivos se hace generalmente después de la fermentación alcohólica (inoculación secuencial) o, más recientemente, junto con las levaduras para inducir fermentaciones alcohólicas y malolácticas concurrentes (coinoculación).
Se sabe que las bacterias malolácticas (ML) pueden influir en los aromas mantecosos del vino debido a la producción de diacetilo, y que se pueden utilizar ciertas técnicas de vinificación para manipular la concentración en el vino después de la FML (Bartowsky y Henschke, 2004). Diversos estudios recientes han destacado el efecto de O. oeni en grupos de compuestos volátiles derivados de la fermentación (ácidos grasos y ésteres de acetatos, ácidos y alcoholes superiores, ácidos orgánicos y aminoácidos). Muchos de estos compuestos volátiles contribuyen a aromas frutales en vinos blancos y tintos (Francis y Newton, 2005); por ejemplo, los ésteres de etil pueden conferirles caracteres afrutados (bayas, piña, plátano), mientras que los ésteres de cadena larga (C6-C10) tienden a dar caracteres jabonosos. Algunos de nuestros estudios en vinos de chardonnay y cabernet sauvignon han mostrado que la FML puede modificar el perfil de ésteres del caldo, con concentraciones de ésteres de etil y de acetato con tendencia a aumentar y disminuir, respectivamente, después de la FML (Bartowsky et al., 2008).
Los vinos tintos con cuerpo medio o carnoso suelen ser descritos con notas intensas de grosella, cereza, frambuesa o ciruela (Iland et al., 2009). Uno de los principales retos para los enólogos es acentuar estos caracteres frutales y minimizar su pérdida durante el proceso de vinificación. Las investigaciones más recientes sugieren que el aroma de bayas en los vinos tintos viene de una compleja interacción entre ésteres frutales, norisoprenoides, dimetilo sulfuro, etanol y otros componentes. Se han identificado algunos grupos de ésteres que contribuyen especialmente a los aromas de bayas rojas y negras (Escudero et al., 2007; Pineau et al., 2009). Basándonos en estos análisis es posible conocer mejor cómo el metabolismo de O. oeni durante la FML puede modificar y acentuar los caracteres frutales, especialmente en los vinos tintos. Se presentan diversos casos en los que se destacan los factores que influyen en la naturaleza y la medida de esta actividad metabólica secundaria asociada con la FML, incluyendo la cepa bacteriana, el momento de la inoculación o los cultivos bacterianos iniciadores, la composición de la matriz vínica y el origen de las uvas.
Influencia de la FML sobre los ésteres frutales del vino y sus propiedades sensoriales
Efecto del régimen de inoculación de la FML respecto a la fermentación alcohólica
Hemos examinado la composición química de mostos y vinos blancos y tintos inoculados, ya sea simultánea o secuencialmente respecto a la fermentación alcohólica, utilizando diferentes cepas comerciales de O. oeni. Pruebas a pequeña escala en vinos de chardonnay y cabernet sauvignon demostraban que la concentración de ésteres totales aumentaba después de la FML, con un mayor incremento cuando las bacterias eran coinoculadas con levaduras al principio de la fermentación alcohólica. La concentración de alcoholes superiores totales disminuía con el régimen de coinoculación de levaduras y bacterias, y se percibían cambios mínimos cuando las bacterias eran inoculadas secuencialmente después de la fermentación alcohólica. Los análisis sensoriales descriptivos de vinos cabernet sauvignon nos han permitido relacionar análisis de composición con cambios aromáticos y sensoriales dados por la FML inducida por bacterias (Bartowsky et al., 2008).
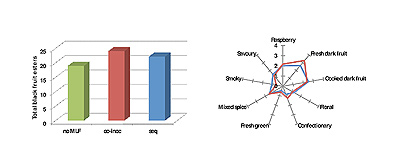
La figura 1 muestra un ejemplo de cómo la coinoculación de O. oeni con levaduras en vinos cabernet sauvignon puede influir en la composición de ésteres y en el perfil sensorial del vino. Este ensayo se llevó a cabo en una bodega sudafricana, a una escala de 1.000 L con triples fermentaciones. Los vinos coinoculados con levaduras y bacterias se describen con «notas afrutadas» y aromas «frescos de frutos negros». Generalmente, la concentración total de ésteres de frutos negros aumentaba con la FML, pero se observaban concentraciones más elevadas en vinos coinoculados, hecho que relaciona la composición química con la descripción sensorial y aromática.
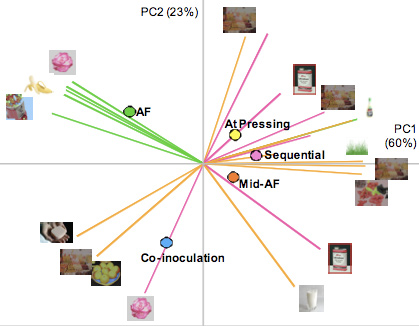
Un estudio reciente investiga los efectos de inocular uvas syrah con bacterias ML en diferentes momentos de la fermentación alcohólica (en el inicio [coinoculació], en la mitad de la fermentación alcohólica, durante el prensado y después de la fermentación alcohólica [secuencial]). Y demostraba que la coinoculación reduce el tiempo total de vinificación (fermentaciones alcohólica + maloláctica) en unas seis semanas: los vinos coinoculados lo completan en tres, mientras que con la inoculación secuencial se alarga hasta nueve (Abrahamse y Bartowsky, 2011). La representación gráfica del análisis de componentes principales (ACP) para los respectivos tratamientos de inoculación y de las composiciones de ésteres volátiles (ésteres de etilos grasos y de acetato) y de alcoholes superiores (fig. 2) revelaba que el tratamiento de coinoculación producía vinos claramente diferenciados de los obtenidos con los otros tratamientos, y también del vino que solo seguía una fermentación alcohólica; la inoculación bacteriana en momentos posteriores dentro del proceso de la fermentación alcohólica o después de esta producía vinos muy similares entre ellos. Estos datos demostraban claramente que los vinos obtenidos con tratamientos de coinoculación tenían perfiles muy diferentes a los volátiles afrutados.
Efecto del origen de la uva, la cepa bacteriana maloláctica y la composición del vino
Se llevaron a cabo estudios con cabernet sauvignon australiano para determinar el efecto de la variación en la cepa de O. oeni sobre la síntesis de ésteres que contribuyen a aromas afrutados, así como la importancia de la composición del vino antes de la FML y de la región vitivinícola (origen del cabernet sauvignon). Se determinaron algunos de los ésteres propuestos por Escudero et al., (2007) y Pineau et al., (2009), y se utilizaron conjuntamente como parámetros químicos para medir los potenciales caracteres sensoriales de tipo frutal y bayas.
El metabolismo de los compuestos del vino producidos por O. oeni vendrán determinados por las diferencias genéticas entre las cepas y la concentración de metabolitos en el vino. Se conoce bien la variabilidad en la producción de diacetilo, que confiere las notas mantecosas al vino. De manera similar, las figuras 3 y 4 muestran que hay variabilidad en la capacidad de producir ésteres entre cepas de O. oeni.
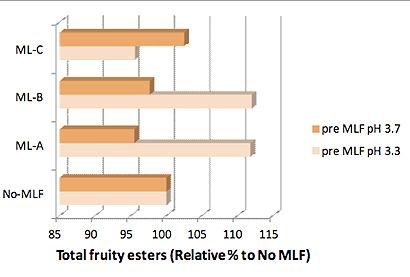
La composición química del vino, incluyendo las concentraciones de alcohol y de SO2 y el pH, tienen un efecto importante en la capacidad de crecimiento de las cepas de O. oeni y en si pueden completar la FML. El pH del vino es también un factor importante sobre la actividad metabólica de O. oeni. Por ejemplo, esta especie tendrá más preferencia por metabolizar azúcares que no ácidos orgánicos a pH elevados (Krieger et al., 2000). De manera similar, nuestros estudios demuestran que, antes de la FML, el pH del vino puede influir en las concentraciones resultantes de compuestos volátiles derivados de la fermentación. Para dos o tres de las cepas bacterianas probadas (ML-A y ML-B), la concentración total de ésteres frutales era mucho más elevada en muestras de vinos a pH 3,3 (tomadas antes de la fermentación) que en muestras de vino que no habían seguido FML. Y al revés, a pH más elevados (3,7) y antes de la FML, estas dos cepas disminuyen la concentración de ésteres frutales totales respecto a las muestras de vinos sin FML (fig. 3). En contraste, este efecto del pH pre-FML se revertía en cierta medida con la cepa ML-C.
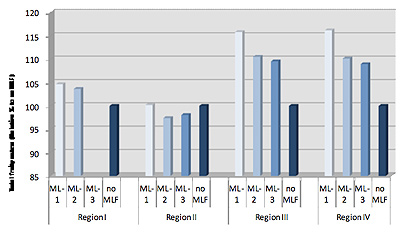
Otro factor que contribuirá al contenido de los ésteres frutales que producirán las cepas de O. oeni es la composición en precursores iniciales del vino. Hemos estudiado este parámetro utilizando uvas de cabernet sauvignon procedentes de diferentes regiones vitivinícolas del sur de Australia (llamadas I, II, III y IV). Antes de la FML, el contenido alcohólico de los vinos era similar (13,9%, 14,7%, 14,4% y 14,0%, respectivamente) y se ajustaban todos a un pH de 3,45. Se inducía la FML con tres cepas de O. oeni. Las cepas ML-1 y -2 metabolizaban completamente el ácido málico en unos 20-25 días, mientras que la cepa ML-3 tardaba un poco más en completar la FML (25-37 días). No la completaba totalmente con el vino de la región I.
Agradecimientos
La figura 4 muestra la producción de ésteres frutales totales para tres cepas de O. Oeni en los cuatro vinos cabernet sauvignon. Las regiones III y IV mostraban un aumento relativamente mayor de ésteres frutales, mientras que la región II se veía poco afectada por la FML. Esta variación en los efectos de la FML sobre la concentración de ésteres entre diferentes regiones refleja muy probablemente oscilaciones regionales en el contenido de precursores.
Bibliografía
. Abrahamse C.E.; Bartowsky, E.J. Timing of malolactic fermentation inoculation in Shiraz grape must and wine: Influence on chemical composition. World Journal of Microbiology & Biotechnology 2011. DOI: 10.1007/s11274-011-0814-3.
. Bartowsky, E., Costello, P.; McCarthy, J. (2008) MLF – adding an ‘extra dimension’ to wine flavour and quality. Australian & New Zealand Grapegrower & Winemaker 2011; 533a: 60-5.
. Bartowsky, E.J.; Henschke, P.A. The ‘buttery’ attribute of wine – diacetyl – desirability, spoilage and beyond. International Journal of Food Microbiology 2004; 96: 235-52.
. Bartowsky, E.J.; Pretorius, I.S. Microbial formation and modification of flavour and off-flavour compounds in wine. In: Biology of microorganisms on grapes, in must and wine. H. König, G. Unden y J. Fröhlich (eds.) Heidelberg: Springer, 2008: 211-33.
. Escudero, A.; Campo, E.; Farina, L.; Cacho, J.; Ferreira, V. Analytical characterization of the aroma of five premium red wines. Insights into the role of odor families and the concept of fruitiness of wines. Journal of Agricultural and Food Chemistry 2007; 55: 4501-10.
. Francis, I.L.; Newton, J.L. Determining wine aroma from compositional data. Australian Journal of Grape and Wine Research 2005; 11: 114-26.
. Iland, P.; Gago, P.; Caillard, A.; Dry, P. A taste of the world of wine. Campbelltown, South Australia: Patrick Iland Wine Promotions Pty Ltd., 2009.
. Krieger, S.A.; Lemperle, E.; Ernst, M. Management of malolactic fermentation with regard to flavor modification in wine. Proceedings of 5th International Symposium on Cool Climate Viticulture and Oenology. Melbourne, Australia: Eds. City, 2000.
. Pineau, B.; Barbe, J.C.; Van Leeuwen, C.; Dubourdieu, D. Examples of Perceptive Interactions Involved in Specific “Red-” and “Black-berry” Aromas in Red Wines. Journal of Agricultural and Food Chemistry 2009; 57: 3702-8.
. Swiegers, J.H.; Bartowsky, E.J.; Henschke, P.A.; Pretorius, I.S. Yeast and bacterial modulation of wine aroma and flavour. Australian Journal of Grape and Wine Research 2005; 11: 139-73.



{kind=link}